Edited by Bruce S. McEwen, The Rockefeller University, New York, NY, and approved July 10, 2017 (received for review April 7, 2017)

Nina Marsha, Dirk Scheelea, Justin S. Feinsteinb,c, Holger Gerhardtd, Sabrina Strange, Wolfgang Maier f,g,and René Hurlemanna,f,1
a Division of Medical Psychology, University of Bonn Medical Center, 53105 Bonn, Germany;
b Laureate Institute for Brain Research, Tulsa, OK 74136;cOxleyCollege of Health Sciences, University of Tulsa, Tulsa, OK 74104;
d Center for Economics and Neuroscience, University of Bonn, 53012 Bonn, Germany;
e Department of Psychology, University of Lübeck, 23562 Lübeck, Germany;
f Department of Psychiatry, University of Bonn Medical Center, 53105 Bonn,Germany;
g German Center for Neurodegenerative Diseases, Helmholtz Association, 53175 Bonn, Germany
https://www.pnas.org/doi/epdf/10.1073/pnas.1705853114
Never before have individuals had to adapt to social environmentsdefined by such magnitudes of ethnic diversity and cultural differ-entiation. However, neurobiological evidence informing about strate-gies to reduce xenophobic sentiment and foster altruistic cooperationwith outsiders is scarce. In a series of experiments settled in thecontext of the current refugee crisis, we tested the propensity of 183Caucasian participants to make donations to people in need, half ofwhom were refugees (outgroup)and half of whom were natives(ingroup). Participants scoring low on xenophobic attitudes exhibitedan altruistic preferencefor the outgroup, which further increased afternasal delivery of the neuropeptide oxytocin. In contrast, participantswith higher levels of xenophobia generally failed to exhibit enhancedaltruism toward the outgroup. This tendency was only countered bypairing oxytocin with peer-derivedaltruistic norms, resulting in a 74%increase in refugee-directed donations. Collectively, these findingsreveal the underlying sociobiological conditions associated withoutgroup-directed altruism by showing that charitablesocial cues co-occurring with enhanced activity of the oxytocin system reduce theeffects of xenophobia by facilitating prosocial behavior towardrefugees.
altruism|ingroup|outgroup|oxytocin|refugees
At this time, we are witnessing one of the largest movements ofrefugees since the end of World War II (1, 2). Ongoing conflicts, persecution, and poverty in the Middle East and Africahave continued forced displacement of more than 65 millionpeople since 2015 (2). Accommodating the large influx of migrantsnot only challenges the humanitarian capacities of Europeancountries but also requires their native populations to adjust torapid growths in ethnic diversity, religious pluralism, and culturaldifferentiation. However, the impetus to adapt to changing socialecosystems is susceptible to considerable interindividual hetero-geneity (3). Resistance to this transition often goes along withxenophobic sentiment (4), and as a consequence, recent electionsin Europe have favored populist candidates who have openlyexpressed xenophobic attitudes toward refugees (5). However, atthe same time, volunteer work for migrants in the hosting coun-tries has reached all-time highs and is estimated to exceed1.6 million hours per month in Germany alone (6). In the face ofgrowing tensions over differences in ethnicity, religion, and culture(3), there is an urgent need for devising strategies for helpingfoster the social integration of refugees into Caucasian societies.As a result of evolutionary selective processes, humans possessa genuine propensity to contrast ingroup members (“us”)fromoutgroup members (“them”)(3).Thisdichotomyisadaptive,asingroup members could not have survived and reproduced withoutaltruistic cooperation [i.e., the goodwill and reciprocity of otheringroup members (7, 8)]. Only recently has neuroscience begun todissect the biological components of altruistic cooperation andidentified oxytocin (OXT), an evolutionarily conserved peptidesignaling pathway originating in the mammalian hypothalamus (9),to be a key modulator (10). These insights could be gained becausentranasally administered OXT (OXTIN) penetrates the brain (11–13) and alters measures of neural and behavioral response (14).Specifically, OXTIN has been revealed to enhance social co-operation (15), generosity (16), and empathy (17, 18); to induce analtruistic response bias away from nonsocial toward social priorities(19); and to reinforce parochial preferences for outgroup hostilityand ingroup centricity (20, 21). Consistent with the latter are find-ings from field studies of wild chimpanzees showing that heightenedendogenous release of OXT correlates with greater ingroup co-hesion during intergroup conflict (22). Furthermore, OXTIN facilitates social norm conformity (23–25). Social norms, and personally costly sanctions against defectors of these norms, an inclination defined as altruistic punishment, may have evolved to protect ingroup biases from erosion through selfish motives (26–29). The biblical parable of the Good Samaritan (Luke 10:25–16:17) describes an ethical maxim of helping strangers who have fallen inneed. As such, it not only captures the essence of altruistic behavior by emphasizing the personal costs of selflessness toward others but also represents a formidable example that norm-enforced altruistic cooperation is by no means limited to the ingroup, but can evenextend to outgroup members in ways neither precisely understood nor systematically researched. Here, we hypothesize that normative incentives co-occurring with enhanced activity of the OXT system exert a motivational force for inducing altruism toward strangers even in the most selfish and xenophobic individuals. To specifically test this hypothesis, the present study was devised to examine socialnorms, administered in the presence vs. absence of a norm-enforcing treatment with OXTIN, for their efficacy to promote altruistic re-sponses in subjects scoring high on a xenophobia inventory (Xi).
Experiment 1To this end, the rationale of Experiment 1 was to generatenormative cues for altruistic responding toward refugees, on thebasis of an incentivized donation task framed in the context of Europe’s refugee crisis (Materials and Methods). This paradigmwas composed of 50 authentic case vignettes briefly describingthe personal needs of poor people, half of which were portrayedas refugees (outgroup) and half as natives (ingroup), re-spectively. The personal needs comprised those elements theUnited Nations has defined as minimum standards for leading asafe and dignified life (30); that is, access to food, adequate housing, or participation in social and cultural life (31, 32).
Assignment of cases to ingroup vs. outgroup frames was balancedacross participants to rule out systematic bias. Subjects were endowed with EUR 50 and could donate a maximum of EUR1 to each case, leaving them the rest (EUR 0–50) as personal payoff. Before testing, subjects prejudicial attitudes to ward refugees was assessed by measuring their individual Xiindex (33)(Materials and Methods). In Experiment 1, a total of 76 healthy female (n=53) and male (n=23) undergraduate students (mean age±SD, 21.2±3.0 y) completed the donation task. Forthe purpose of generating an altruistic norm, subjects were as-sembled in a lecture hall, enabling reputation pressures toprompt potential donors to respond more generously. Indeed, results show that participants contributed more than 30% oftheir endowment. Interestingly, the donations devoted to refugees were 19% higher (EUR 8.03±6.74) than those to natives [EUR 6.71±6.86;t(75) = 5.35;P<0.01;d = 0.19].
This bias in dicates an altruistic preference for the outgroup and was lowestin Xihigh scorers (r=−0.34;P<0.01) (Fig. 1A). An additionalanalysis including gender as a between-subject variable showedthat neither the donations to natives and refugees nor the out-group bias (donations to refugees minus donations to natives)was influenced by gender (allP≥0.86).Experiments 2 and 3Experiments 2 and 3 were carried out within the same randomizedcontrolled trial and involved an independent sample of 107 maleparticipants (mean age±SD, 24.1±3.2 y). Before testing, theirprejudicial attitudes toward refugees were measured by the Xiindex (33). Subjects self-administered a 24-IU dose of OXTINor placebo (PLCIN), which was instructed and supervised by ablinded experimenter in accordance with the latest standardiza-tion guidelines (34). Subsequently, subjects were placed alone inseparate test cubicles and tested on the donation task establishedin Experiment 1 (Fig. S1). Based on the subjects’Xiindex, thesample was median-dichotomized, resulting inn=53 Xihighscorers andn=54 Xilow scorers. A repeated-measures analysis ofvariance with the between-subjects factors treatment (OXTIN,PLCIN)andXiindex (low, high), the within-subjects variable“frame”(ingroup, outgroup) and the donated sums as a de-pendent variable yielded main effects of treatment [F(1,103)=4.64;P=0.03;η2=0.04], Xiindex [F(1,103)=13.51;P<0.01;η2=0.12],and frame [F(1,103)=24.70;P<0.01;η2=0.19]. Specifically,OXTINpromoted generosity toward both the outgroup [OXTIN,EUR 4.41±5.73; PLCIN, EUR 2.62±3.00;t(73.90)=2.00;P=0.05;d=0.40] and the ingroup [OXTIN,EUR3.62±5.44; PLCIN,EUR 1.99±2.58;t(69.88)=1.94;P=0.06;d=0.39], evident in a68% (outgroup) and an 81% (ingroup) increase in the donatedsums (Fig. 1B). Furthermore, we detected an interaction of frameand Xiindex [F(1,103)=12.15;P<0.01;η2=0.11]; that is, irre-spective of OXTINtreatment, Xilow scorers’contributions were31% larger for the outgroup than for the ingroup [t(53)=4.99;P<0.01;d=0.22], which replicates the outgroup favoritism observedin Experiment 1. As expected, this outgroup bias was absent in X
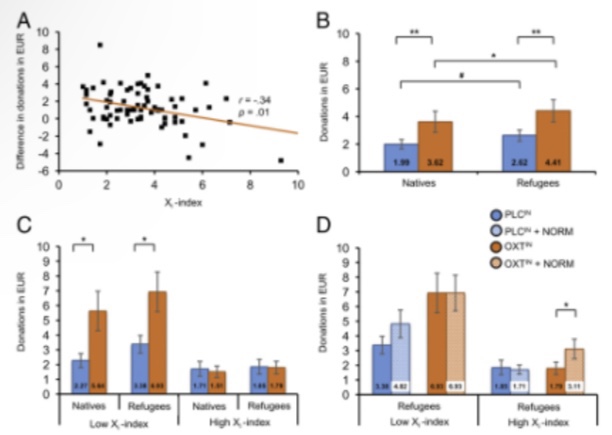
Fig. 1.The sociobiological conditions that reduce the effects of xenophobia by facilitating prosocial behavior toward refugees.(A)In Experiment 1 (n=76),altruistic donations for the outgroup were lowest in those scoring high on the Xiindex (scores ranging from 1=low to 10=high).(B)In Experiment 2, anindependent sample of 107 male participants received OXTIN(n=51) or placebo (n=56) before the donation task. OXTIN promoted a 68% (outgroup) and an81% (ingroup) increase in the donated sums.(C)Based on the subjects’Xiscores from the 7-item scale of the realistic threat inventory, the sample was median-dichotomized (n=54 Xilow scorers;n=53 Xihigh scorers). Xilow scorers who received OXTIN(n=26) more than doubled their donations to both groups,whereas the peptide did not induce generosity in Xihigh scorers (n=25).(D)Pairing OXTIN with peer-derived norms in Experiment 3 prompted Xihigh scorersto increase their outgroup-related donations by 74%. OXTIN+Norm, intranasal oxytocin paired with a peer-derived norm; PLCIN+Norm, intranasal placebopaired with a peer-derived norm. Error bars indicate the SEM. #P=0.056; *P<0.05; **P<0.01
high scorers, whose donations were not significantly different be-tween the two frames [ingroup, outgroup;t(52)=1.43;P=0.16;d=0.09]. An additional interaction of treatment and Xiindex[F(1,103)=5.43;P=0.02;η2=0.05] reflects the inefficacy ofOXTINtreatment alone to induce generosity in Xihigh scorers (allPvalues>0.75), whereas the peptide more than doubled thecontributions of Xilow scorers to both the outgroup [t(34.52)=2.40;P=0.02;d=0.68] and the ingroup [t(31.20)=2.37;P=0.02;d=0.68; Fig. 1C].To address the question of whether administration of a peer-derived altruistic norm in addition to OXTINcould augment al-truistic responses in Xihigh scorers, the task was reiterated inExperiment 3, with the exception that this time, each case pre-sentation also included information about the average contributionchoices of all their male and female peers enrolled in Experiment 1(Fig. S2). A repeated-measures ANOVA with“norm”(present,absent) as an additional within-subject variable revealed a three-way interaction among treatment, Xiindex, and norm [F(1,103)=5.51;P=0.02;η2=0.05]. Not surprisingly, deviation from the normwas relatively low for Xilow scorers due to their distinct altruistictendency displayed in Experiment 2, which is consistent with atrend-to-significant effect of norm administration in PLCIN-treatedsubjects [outgroup,t(27)=1.73 (P=0.095;d=0.34); ingroup,t(27)=1.87 (P=0.07;d=0.35)] and no effect at all in OXTIN-treatedsubjects (allPvalues>0.40). Intriguingly, application of OXTINinconjunction with the altruistic norm prompted Xihigh scorers,who had been resistant to either of these interventions alone (allPvalues>0.64), to increase their outgroup-related donations by 74%[EUR 3.11±3.37 with norm vs. EUR 1.79±2.14 without norm;t(24)=2.61;P=0.02;d=0.47; Fig. 1D]. This effect was weaker forthe ingroup [EUR 2.46±2.93 with vs. EUR 1.51±2.00 withoutnorm;t(24)=1.81;P=0.08;d=0.38]. The facilitation of altruismobserved in Xihigh scorers becomes even more obvious when rel-ativizing the donated sums to those obtained in Experiment 1; rel-ative to this 100% benchmark, donations in Xihigh scorers climbedfrom 23% to 38% as a consequence of pairing OXTINtreatmentwith the peer-derived altruistic norm.DiscussionAs mentioned earlier, the parable of the Good Samaritan describesa highly influential ethical maxim of helping outgroup members whohave fallen in need, and has attained paramount political signifi-cance in the African-American civil rights movement (35). However,as yet, the social and biological conditions promoting such outgroup-directed altruism have not been determined. Here, we show thatenhanced activity of the OXT system paired with charitable socialcues can help counter the effects of xenophobia by fostering altruismtoward refugees. These results areespecially important in the lightof evidence that even a minority of selfish noncooperators (Fig. 1C)may suffice to force the majority ofaltruists to defect, resulting in arapid decay of altruistic cooperation within a population (7).Therefore, selfish motives impose an impending threat to altruisticcooperation (27). Here, we demonstrate that normative incentivesco-occurring with elevated activity of the OXT system exert a mo-tivational force for inducing altruistic cooperation with outsiders,even in those individuals who refuse to do so in the absence of suchexogenous triggers. Since their selfless behavior only emerged as aresult of OXT-enhanced social norm compliance, it is extrinsicallymotivated. However, even intrinsically motivated (i.e., self-generated) forms of altruism may build on internalized social (e.g.,parental) norms and engage endogenous OXT signaling. Consistentwith previous observations that OXTINincreases generosity per se(16, 36), we found a generalized increase in donations toward boththe outgroup and the ingroup (Fig. 1B). However, this effect wasmainly driven by the higher donations of the Xilow scorers (Fig.1C), and therefore is in line with evidence emphasizing a sensitivityof OXTINeffects to person- and context-dependent factors (19, 37,38). Whereas previous studies have focused either on the efficacy ofingroup norms as a potential means of stabilizing altruistic co-operation (28) or on the facilitating effects of OXT signaling onsocial conformity (23, 24), none have combined both interventionsto enhance social norm adherence. Here, we provide evidence that axenophobic rejection of refugeescan be reversed by coupling en-hanced activity of the OXT system to a normative incentive forcooperation with peers; neither intervention alone was sufficient toalter selfish responses in Xihigh scorers, illustrating the relativeresistance of outgroup rejection to exogenous modification. Unfortunately, open and latent xenophobia continue to be amajor challenge for European democracies. Since foraging socie-ties were afflicted by intergroup conflict at all times, a strong in-clination to categorically differentiate between ingroup (“us”)andoutgroup (“them”) members may have conferred evolutionaryadvantages (39). Warfare may even have catalyzed cultural selec-tion, as the dominant groups have forced their social preferencesand norms on the defeated groups (7, 27). Our results raise thequestion of whether higher levels of xenophobia could be associ-ated with a reduced sensitivity to others’distress, irrespective oftheir group membership. This may explain why the Xihigh scorersin our sample did not exhibit an altruism bias to either group. Itshould be emphasized, though, that the measured Xiscores rep-resent relatively typical levels of xenophobia within the generalpopulation. Based on the heterogeneity of xenophobic attitudeswithin certain groups and regions (40, 41), future research with afocus on the extreme ends of xenophobia is needed to provide amore nuanced understanding of the effects of OXTINon outgroup-directed altruism. Furthermore, given that OXTINcan producesexual-dimorphic effects (42, 43), we cannot extrapolate our find-ings to women. Clearly, future studies are warranted to explore therelationship between the effects of OXTINon outgroup-directedaltruism and a wider array of person- and context-dependent fac-tors, including sex, age, and self-report measures of xenophobia.
In contrast to previous studies, which have used OXTINto illus-trate a contribution of OXT signaling in parochial altruism, especially under circumstances of intergroup conflict (20, 21), we demonstratethat enhanced activity of the OXT system facilitates social normcompliance, thus inducing altruism toward outgroup members evenin the most selfish and xenophobic individuals. Given evidence in-dicating that social group activities with peers, such as singing in achoir (44), are associated with elevated endogenous OXT release(45), our findings suggest that greater focus should be placed onenabling positive social encounters among citizens of hosting countries that communicate a prosocial norm; that is, by affirming and emphasizing the benefits of ethnic diversity, religious pluralism, and cultural differentiation. This may include the promotion of balanced and informed media reporting, the integration of refugee themes to the curricula of schools and universities, or the organization of events that involve the general public and bring communities together by promoting sustained experience- and information-sharing on thesituation of refugees (46). The effect of solutions combining selectiveenhancement of OXT signaling and peer influence would beexpected to diminish selfish motives, and thereby increase the ease by which people adapt to rapidly changing social ecosystems. More generally, our results imply that an OXT-enforced social norm ad-herence could be instrumental in motivating a more generalized acceptance toward ethnic diversity, religious plurality, and cultural differentiation resulting from migration by proposing that interventions to increase altruism are most effective when charitable social cues instill the notion that one’s ingroup shows strong affection for anoutgroup. Further more, UNESCO has emphasized the importance of developing neurobiologically informed strategies for reducing xe-nophobic, hostile, and discriminatory attitudes (47). Thus, considering OXT-enforced normative incentives in developing future interventions and policy programs intended to reduce outgroup re-jection may be an important step toward making the principle of social inclusion a daily reality in our societies, subjects were assembled in a lecture hall. Using a paper–pencil version of the altruistic donation task, the participants were instructed to indicate the exact amount of their donation for each case vignette on the forms, which werelater collected by the experimenter. By design, this experimental setting en-abled reputation pressures due to social interactions among peers, such thatthe sums donated in Experiment 1 were higher than those from Experiments2 and 3, in which we eliminated social interaction.
Experiments 2 and 3.
For Experiments 2 and 3, a new group of participants wasinvited to sign up for the experiment via the online database hroot (50) of theBonnEconLab. A total of 127 healthy male volunteers enrolled in the double-blind, randomized, parallel-group trial design and self-administered a dose of24 IU (three puffs per nostril, each with 4 IU; Novartis) of OXTINor PLCIN40–45 min before the start of the donation task. The placebo solution containedthe identical ingredients except the peptide itself. The study comprised ascreening session in the morning and an experimental session in the after-noon. Screening entailed the exclusion of current or past physical or psychiatricillness (including drug and alcohol abuse), as assessed by medical history andthe Mini-International Neuropsychiatric Interview (51). To control for possiblepretreatment differences, we assessed anxiety traits with the State-TraitAnxiety Inventory (52), depressive symptoms with the Beck Depression In-ventory (53), early social adversity with the Childhood-Trauma-Questionnaire(54), and autistic-like traits with the Autism-Spectrum-Quotient (55). Further-more, we assessed cooperative and altruistic attitudes based on the subjects’social value orientation (56), as well as empathy with the Interpersonal Re-activity Index (57) and their susceptibility to external (dis)approval with theSocial Desirability Scale (58). In addition, the subjects were asked to indicatetheir personal income and donation behavior during the last year. There wereno a priori differences between the OXTINand PLCINgroups for Xilow andhigh scorers (Tables 2 and 3). Moreover, subjects were naive to prescription-strength psychoactive medication and had not taken any over-the-counterpsychoactive medication in the preceding 4 wk. Participants were asked tomaintain their regular sleep and waking times and to abstain from caffeineand alcohol intake on the day of the test session. During testing, a total of20 participants (OXTIN:n=10; PLCIN:n=10) could not complete the donationtask due to technical issues. Thus, the final analyses were performed on datafrom 107 participants (mean age±SD, 24.16±3.2 y).During Experiments 2 and 3, each participant was seated alone in front of acomputer screen in a separate test cubicle thatwas closed off with curtains, whilebeing exposed to the same donation task and instructional primes as establishedin Experiment 1. This experimental setting eliminated any interactions withpeers or the experimenter. To exclude potential confounding influences of sexdifferences, we compared OXT effects between male participants, rather thanbetween men and women. Participants performed two runs of the donationtask. In Experiment 2, participants had no further information than that theycould donate some, none, or all of their money with a maximum of EUR 1 foreach case vignette (Fig. S1). In Experiment 3, the same sample of participantswas additionally informed about the higher average donations made by theirpeersineachscenario(e.g.,“Your peers previously donated EUR 0.50”)(Fig.S2). The scenarios of the donation task were presented with the softwareQualtrics. Participants were asked to indicate their donation by moving a sliderplaced under each scenario. By varying the starting positions of the slider, eachchoice required a similar effort.Xenophobia index.In a separate screening session, we evaluated xenophobiaby measuring the attitudes toward refugees based on an adapted assessmentinstrument developed by Schweitzer and colleagues (33). Adaptions encom-passed the wording; for example,“Australian refugee”was replaced by“German refugee.”The assessment instrument contained two inventories,in which participants indicated how strongly they associate refugees withrealistic and symbolic threats. In our analyses, we focused on the realisticthreat inventory, which includes seven items and has good internal consis-tency (Cronbach’sα=0.83). This decision was based on previous evidenceindicating that realistic threats (e.g., perceived threats to economic interest,social status, or health), rather than symbolic threats (e.g., perceived threatsto morals, values, or beliefs), more robustly predict perceptions of xeno-phobia (40, 41, 59). The realistic threat scale items encompass differentthreat perceptions; for example:“Refugees are not displacing Germanworkers from their jobs”or“Refugees have increased the tax burden onGermans.”Responses were coded on a 10-point Likert scale, ranging from 1(“I strongly disagree”)to10(“I strongly agree”). All items were recoded suchthat higher values reflected greater feelings of perceived realistic threats.The term Xiindex, which we used for subsequent analyses, refers to a sub-ject’s mean score achieved on the realistic threat inventory.Statistical Analysis.Demographic, neuropsychological, and behavioral datawere analyzed using IBM SPSS Statistics 24. Quantitative behavioral data werecompared using mixed analysis of variance and post hoct-tests. Pearson’sproduct-moment correlation was used for correlation analysis. Eta-squaredand Cohen’sdwere calculated as measures of effect size. For qualitativevariables, Pearson’sχ2tests were used. All reportedPvalues are two-tailed ifnot otherwise stated, andPvalues ofP<0.05 were considered significant.All relevant data are stored on a server of the Division of Medical Psy-chology at the University of Bonn Medical Center and are available for re-search purposes on request by contacting R.H.ACKNOWLEDGMENTS.The authors thank two anonymous referees for theirgenerous comments. This research was supported by a grant from the German Science Foundation (to R.H. and D.S.; HU 1202/4-1 and BE 5465/2-1



Materials and Methods
Our study was approved by the Institutional Review Board of the Faculty of Medicine of the University of Bonn, and was carried out in compliance with the latest revision of the Declaration of Helsinki. All participants gave written, informed consent xperimental Design.The altruistic donation task, which was framed in thecontext of Europe’s refugee crisis, included 50 authentic case vignettes in total.Each of the 25 ingroup case vignettes had a corresponding outgroup coun-terpart (Fig. S1). The descriptions presented in the vignettes were composed ina standardized manner to keep them identical regarding formal criteria such asstyle, word count, and format. Each individual was introduced by his/her name,age, place of birth, country of origin, and personal need. The selection of homecountries for the outgroup frame was based on reports published in 2015 by the European Commission, documenting conflicts, persecution, and povertyin the Middle East and Eastern Europe as the major drivers of forced migrationto the European Union (48, 49). Personal needs comprised those essential ele-ments the United Nations has defined as minimum standards for leading a safe and dignified life (30); that is, access to food, adequate housing, or participationin social and cultural life (31, 32). All 50 case scenarios were balanced in terms of age and sex of the needy persons and were presented in random order. The participants read and completed the donation task at their own pace. Subjectswere endowed with EUR 50 and could donate a maximum of EUR 1 to eachcase, leaving them the rest (EUR 0–50) as personal payoff, if the lottery ticket,which they drew on completion of the donation task, had a number on theinside (chance of winning: 10%). Furthermore, subjects were informed that alltheir donations were subtracted from this endowment of EUR 50.Procedure and Participants.Experiment 1.In Experiment 1, a total of 76 healthy female (n=53) and male(n=23) undergraduate students (mean age±SD, 21.2±3.0 y; Table 1) completed the donation task. The rationale of Experiment 1 was to generate normative cues for altruistic responding toward refugees. For this purpose
Significance
In the midst of rapid globalization, the peaceful coexistence ofcultures requires a deeper understanding of the forces thatcompel prosocial behavior and thwart xenophobia. Yet, theconditions promoting such outgroup-directed altruism have notbeen determined. Here we report the results of a double-blind, placebo-controlled experiment showing that enhanced activityof the oxytocin system paired with charitable social cues canhelp counter the effects of xenophobia by fostering altruismtoward refugees. These findings suggest that the combination of oxytocin and peer-derived altruistic norms reduces outgrouprejection even in the most selfish and xenophobic individuals, and thereby would be expected to increase the ease by whichpeople adapt to rapidly changing social ecosystems.
Author contributions: N.M., D.S., and R.H. designed research; N.M., H.G., and S.S. per-formed research; N.M., D.S., and R.H. analyzed data; and N.M., D.S., J.S.F., H.G., S.S., W.M.,
- DePillis L, Saluja K, Lu D (December 21, 2015) A visual guide to 75 years of major refugeecrises around the world.Washington Post.Available at https://www.washingtonpost.com/graphics/world/historical-migrant-crisis/. Accessed May 20, 2017.
- United Nations High Commissioner for Refugees (UNHCR) (2016) Global trends:Forced displacement in 2015. Available at www.unhcr.org/576408cd7.pdf. AccessedJuly 26, 2017.
- Crisp RJ, Meleady R (2012) Adapting to a multicultural future.Science336:853–855.4.
- United Nations Educational, Scientific and Cultural Organization (2016) Learning tolive together. Available at www.unesco.org/new/en/social-and-human-sciences/themes/international-migration/glossary/xenophobia/. Accessed March 13, 2017.
- Aisch G, Pearce A, Rousseau B (2016) How far is Europe Swinging to the right?NYTimes. Available at https://www.nytimes.com/interactive/2016/05/22/world/europe/europe-right-wing-austria-hungary.html?_r=0. Accessed March 12, 2017
- Quadbeck E (2016) Freiwillige Flüchtlingshelfer leisten Millionen Stunden.
- RheinischePost.Available at http://www.rp-online.de/panorama/deutschland/freiwillige-fluechtlingshelfer-leisten-millionen-stunden-aid-1.5789308. Accessed March 12, 2017.7. Fehr E, Fischbacher U (2003) The nature of human altruism.Nature425:785–791.8. Nowak MA, Sigmund K (2005) Evolution of indirect reciprocity.Nature437:1291–1298.9. Donaldson ZR, Young LJ (2008) Oxytocin, vasopressin, and the neurogenetics of so-ciality.Science322:900–904.10. Hurlemann R, Marsh N (2016) [New insights into the neuroscience of human altruism].Nervenarzt87:1131–1135.11. Born J, et al. (2002) Sniffing neuropeptides: A transnasal approach to the humanbrain.Nat Neurosci5:514–516.12. Lee MR, et al. (March 14, 2017) Oxytocin by intranasal and intravenous routes reachesthe cerebrospinal fluid in rhesus macaques: Determination using a novel oxytocinassay.Mol Psychiatry, 10.1038/mp.2017.27.13. Striepens N, et al. (2013) Elevated cerebrospinal fluid and blood concentrations ofoxytocin following its intranasal administration in humans.Sci Rep3:3440.14. Meyer-Lindenberg A, Domes G, Kirsch P, Heinrichs M (2011) Oxytocin and vasopressinin the human brain: Social neuropeptides for translational medicine.Nat Rev Neurosci12:524–538
Why have we nothing learned from history?

Hinterlasse einen Kommentar